溶酶体
一、形态结构和化学组成
(一)高度异质性的膜性细胞器
溶酶体在其形态大小、数量分布、生理生化性质等各方面都表现出了高度的异质性(heterogenous)。
- 大小差异显著。
- 典型的动物细胞中约含有几百个溶酶体,但是,在不同细胞中溶酶体的数量差异是巨大的。
- 一般而言,在溶酶体中可含有60多种能够分解机体中几乎所有生物活性物质的酸性水解酶,这些酶作用的最适H通常在 3.5~5.5。但是,在每一个溶酶体中所含有的酶的种类却是有限的。
(二)共同特征是含有酸性水解酶
尽管溶酶体是一种具有高度异质性的细胞器,但是却也具有许多重要的共性特征:
- 都是由一层单位膜包裹而成的囊球状结构小体;
- 均含有丰富的酸性水解酶,包括蛋白酶、核酸酶、脂酶、糖苷酶、磷酸酶和溶菌酶等多种酶类。其中,酸性磷酸酶是溶酶体的标志酶;
- 溶酶体膜中富含两种高度糖基化的穿膜整合蛋白IgpA和IgpB。它们分布在溶酶体膜腔面,可能有利于防止溶酶体所含的酸性水解酶对其自身膜结构的消化分解;
- 溶酶体膜上嵌有V-型质子泵,可依赖水解ATP释放出的能量将H+逆浓度梯度地泵入溶酶体中,以形成和维持溶酶体囊腔中酸性的内环境。
(三)溶酶体膜糖蛋白家族具有高度同源性
溶酶体膜糖蛋白家族——溶酶体结合膜蛋白(lysosomal--asso ciated membrane protein,LAMP)或溶酶体整合膜蛋白(lysosomal integral membrane protein,LIMP)。该类蛋白的肽链组成结构包括:一个较短的N-端信号肽序列、一个高度糖基化的腔内区、一个单次跨膜区和一个由10个左右的氨基酸残基组成的C-端胞质尾区。
- 在寡糖链的末端均含有唾液酸。这种高度的糖基化使得蛋白质的等电点极低而呈酸性。
- 溶酶体整合膜蛋白高度保守的C-端胞质尾区区段可能是该类蛋白质从高尔基复合体向溶酶体运输的通用识别信号。这也提示:N-糖基化并非是这些蛋白质必需的转运信号。
二、形成与成熟过程
溶酶体的形成是一个由内质网和高尔基复合体共同参与,集胞内物质合成加工、包装、运输及结构转化为一体的复杂而有序的过程。就目前的普遍认识,以溶酶体酶蛋白在附着型多聚核糖体上的合成为起始,溶酶体的形成主要经历以下几个大的阶段:
1.酶蛋白的N-糖基化与内质网转运
合成的酶蛋白前体进入内质网网腔,经过加工、修饰,形成N-连接的甘露糖糖蛋白;再被内质网以出芽的形式包裹形成膜性小泡,转送运输到高尔基复合体的形成面。
2.酶蛋白在高尔基复合体内的加工与转移
在高尔基复合体形成面囊腔内磷酸转移酶与N-乙酰葡萄糖胺磷酸糖苷酶的催化下,寡糖链上的甘露糖残基磷酸化形成甘露糖-6-磷酸(mannose-6-phosphate,M-6-P),此乃溶酶体水解酶分选的重要识别信号。 #?
3.酶蛋白的分选与转运
当带有M-6-P标记的溶酶体水解酶前体到达高尔基复合体成熟面时,被高尔基复合体网膜囊腔面的受体蛋白所识别、结合,随即触发高尔基复合体局部出芽和网膜外胞质面网格蛋白的组装,并最终以表面覆有网格蛋白的有被小泡(coated vesicle)形式与高尔基复合体囊膜断离。以M-6-P为标志的溶酶体酶分选机制是目前了解比较清楚的一条途径,但并非溶酶体酶分选的唯一途径。
4.内体性溶酶体的形成与成熟
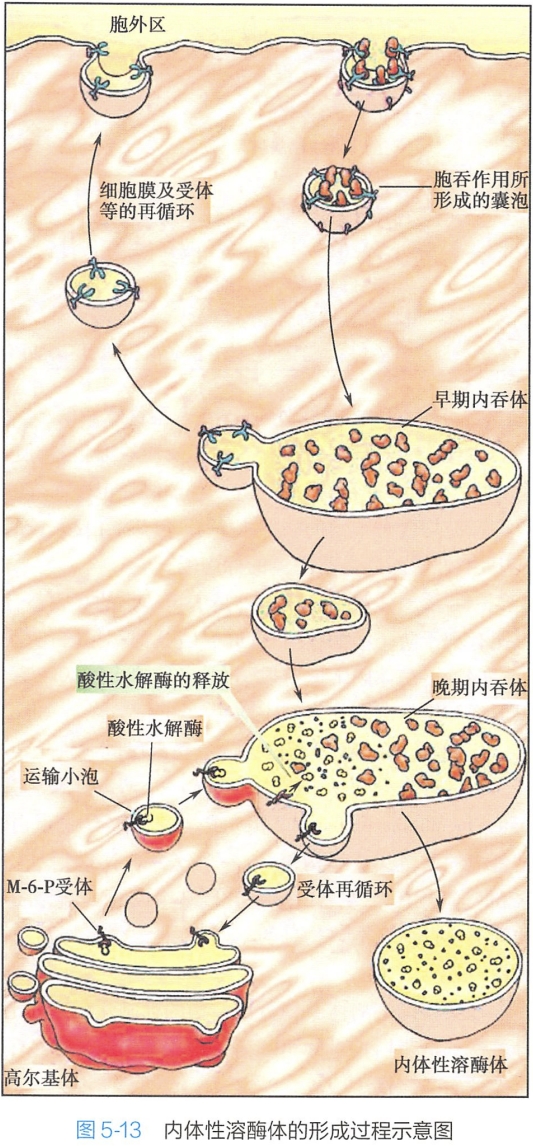
有被小泡很快脱去网格蛋白外被形成无被小泡,与晚期内体(也称晚期内吞体)融合,形成内体性溶酶体。
- 内体(endosome),是由细胞的胞吞作用形成的一类异质性脱衣被膜泡,依其发生阶段而分为早期内体(early endosome)和晚期内体。
- 早期内体是指经由胞吞作用入胞后最初的脱衣被膜泡,其囊腔中含有胞吞物质,是一个H+和细胞外液大致相当的碱性内环境。
- 当早期内体通过分拣、分离出带有质膜受体的再循环内体(recycling endosome)后,即完成了从早期内体向晚期内体的转化,而再循环内体则返回并重新融入到质膜中。( #? 前文LDL这里是pH降低后才分离,当然,不一样确实有可能,但是前文的所谓融合多半是错误的)
- 晚期内体相对靠近于细胞核一侧,和来源于高尔基复合体的那些含有酸性水解酶的运输小泡融合之后,在其囊膜上质子泵的作用下,将胞质中的H+泵入,使其腔内pH从7.4左右下降到6.0以下。在改变了的酸性内环境条件下,溶酶体酶前体从与之结合的M-6-P膜受体上解离,并通过去磷酸化而成熟;同时,膜M-6-P受体则出芽形成运输小泡,重新回到高尔基复合体成熟面的网膜上。经历这些系列变化之后,最终形成内体性溶酶体。
三、类型
(一)以功能状态区分
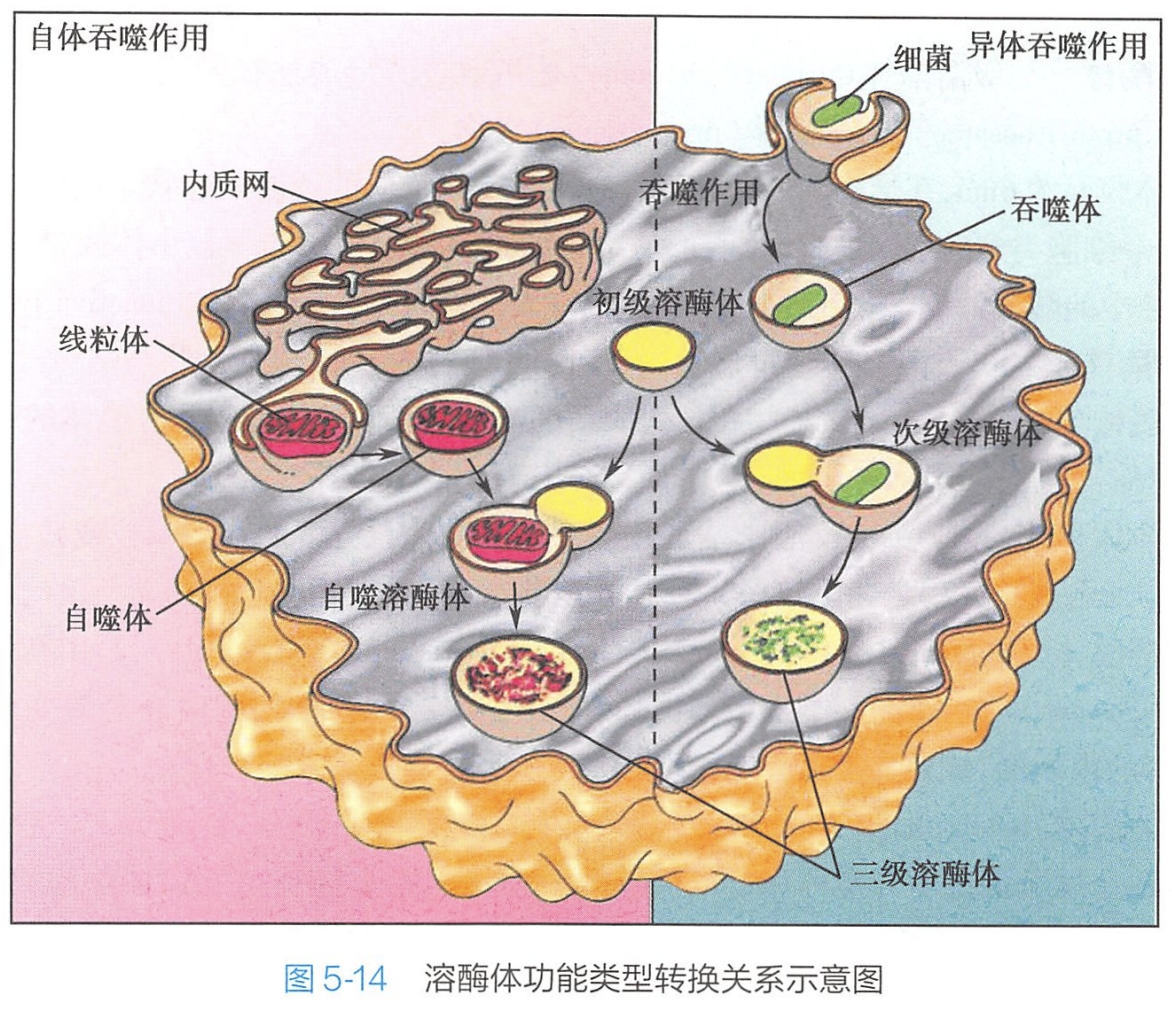
1.初级溶酶体/原溶酶体/前溶酶体/无活性溶酶体
初级溶酶体(primary lysosome)是指通过其形成途径刚刚产生的溶酶体。初级溶酶体囊腔中的酶通常处于非活性状态,因此也有人称之为无活性溶酶体(inactive lysosome)。(大概就是有被运输小泡刚刚与晚期内体融合后的产物阶段)
2.次级溶酶体/消化泡
当初级溶酶体成熟,接受来自细胞内、外的物质,并与之发生相互作用时,即成为次级溶酶体(secondary lysosome)。依据次级溶酶体中所含作用底物之性质和来源的不同,又把次级溶酶体分为不同的类型:
- [200 学习/201 细胞生物学/第16章 细胞死亡/第3节 细胞自噬/细胞自噬|自噬溶酶体]:又称自体吞噬泡(autophagic vacuole),是由初级溶酶体融合自噬体后形成的一类次级溶酶体,其作用底物主要是细胞内衰老蜕变或残损破碎的细胞器(如损害的内质网、线粒体等)或糖原颗粒等其他胞内物质。 #?
- 异噬溶酶体(heterophagic lysosome):又称异体吞噬泡(heterophagic vacuole),是由初级溶酶体与细胞通过胞吞作用所形成的异噬体(heterophagolysosome,包括吞噬体与吞饮体小泡)相互融合而成的次级溶酶体,其作用底物源于外来异物。
- 吞噬溶酶体(phagolysosome):是由吞噬细胞吞入胞外病原体或其他外来较大的颗粒性异物所形成的吞噬体与初级溶酶体融合而成的次级溶酶体。由于吞噬溶酶体与异噬溶酶体的作用底物均为细胞外来物,因此,二者之间并无本质上的区别。
3.三级溶酶体/后溶酶体/终末溶酶体
次级溶酶体在完成对绝大部分作用底物的消化、分解作用之后,尚会有一些不能被消化、分解的物质残留于其中,随着酶活性的逐渐降低以至最终消失,进入了溶酶体生理功能作用的终末状态。此时又被易名为残余体(residual body)。这些残余体,有些可通过细胞的排遗作用,以胞吐的方式被清除、释放到细胞外去;有些则可能会沉积于细胞内而不被外排。例如:
- 常见于脊椎动物和人类神经细胞、肝细胞、心肌细胞内的脂褐质(lipofuscin)。脂褐质是由单位膜包裹的非规则形态小体,内含脂滴和电子密度不等的深色调物质。
- 含铁小体(siderosome)。含铁小体内部充满电子密度较高的含铁颗粒。当机体摄入大量铁质时,在肝、肾等器官组织的巨噬细胞中常会出现许多含铁小体。
- 见于肿瘤细胞、某些病毒感染细胞、大肺泡细胞和单核吞噬细胞中的髓样结构(myelin figure)。其最显著的特征是内含板层状、指纹状或同心层状排列的膜性物质。和髓样体不同。
(二)以形成过程区分
1.内体性溶酶体(endolysosome,也称内溶酶体)
由高尔基复合体芽生的运输小泡并入经由细胞胞吞(饮)作用形成的晚期内体(late endosome)形成。比如初级溶酶体
2.吞噬性溶酶体
由内体性溶酶体和自噬体或异噬体相互融合而成
四、功能
(一)分解外来物质及清除衰老、残损的细胞器
溶酶体能够通过形成异噬溶酶体和自噬溶酶体的不同途径,及时地对经胞吞(饮)作用摄入的外来物质或细胞内衰老、残损的细胞器进行消化,使之分解成为可被细胞重新利用的小分子物质,并透过溶酶体膜释放到细胞质基质,参与细胞的物质代谢。
(二)溶酶体具有物质消化与细胞营养功能
溶酶体作为细胞内具有消化功能的细胞器,在细胞饥饿状态下,可通过分解细胞内的一些对于细胞生存并非必需的生物大分子物质,为细胞的生命活动提供营养和能量,维持细胞的基本生存。
(三)溶酶体是机体防御保护功能的组成部分
细胞防御是机体免疫防御系统的重要组成部分,而溶酶体强大的物质消化和分解能力则是细胞实现其免疫防御功能的基本保证和基本机制。通常,在巨噬细胞中均具有发达的溶酶体,被吞噬的细菌或病毒颗粒,最终都是在溶酶体的作用下而得以杀灭,并被分解消化的。
(四)溶酶体参与某些腺体组织细胞分泌过程的调节
溶酶体常常在某些腺体组织细胞的分泌活动过程中发挥着重要的作用。例如,储存于甲状腺腺体内腔中的甲状腺球蛋白,首先要通过吞噬作用进入分泌细胞内,在溶酶体中水解成甲状腺素,然后才被分泌到细胞外。
(五)溶酶体在生物个体发生与发育过程中起重要作用
在动物精子中,溶酶体特化为其头部最前端的顶体(acrosome),当精子与卵子相遇、识别、接触时,精子释放顶体中的水解酶,溶解、消化围绕卵细胞的滤泡细胞及卵细胞外被,从而为精核的入卵受精打开一条通道。在无尾两栖类动物个体的变态发育过程中,其幼体尾巴的退化、吸收;脊椎动物生长发育过程中骨组织的发生及骨质的更新;哺乳动物子宫内膜的周期性萎缩、断乳后乳腺的退行性变化、衰老红细胞的清除以及某些特定的细胞编程性死亡等,都离不开溶酶体的作用。